Connaître l’épidémiologie et les caractéristiques de l’hépatite E en France
Connaître les méthodes diagnostiques virologiques : quels tests dans quelle situation clinique
Connaître l’éventuel passage à la chronicité et son traitement
Connaître la prévention
Vidéo
Introduction
L’existence d’un agent viral différent du virus de l’hépatite A et responsable d’hépatites à transmission entérique a d’abord été envisagée lors d’une épidémie gigantesque survenue en 1978 au Cachemire. Outre l’absence de marqueurs sérologiques des hépatites A et B, des formes cliniques particulièrement sévères étaient observées chez les femmes enceintes [1]. Ce nouvel agent a ensuite été découvert lors de l’investigation d’une épidémie d’hépatites inexpliquées survenue en 1981 dans un camp militaire soviétique en Afghanistan. Après avoir ingéré un pool d’extrait de selles, un chercheur volontaire a développé une hépatite aiguë et des particules virales sphériques ont pu être visualisées par microscopie électronique dans ses propres selles [2]. C’est seulement en 1990 que le génome viral a pu être partiellement cloné et séquencé à partir de la bile de macaques infectés expérimentalement et que l’agent des hépatites non-A non-B à transmission entérique a été nommé Virus de l’hépatite E (VHE) [3]. L’hépatite E est alors une cause majeure d’hépatite aiguë dans les pays en développement d’Asie et d’Afrique, l’infection étant liée à l’absence ou à la défaillance de l’approvisionnement en eau potable. En 1997, la découverte du VHE porcin aux USA [4] et la description du premier cas autochtone associé à cette souche [5] a soulevé la question du risque zoonotique dans les pays développés. La distribution mondiale de l’hépatite E est maintenant reconnue avec cependant des modes de transmission et des formes cliniques qui diffèrent selon le génotype viral et la zone géographique.
Caractéristiques virologiques et épidémiologie moléculaire
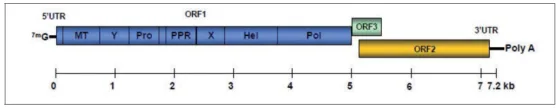
Dans les selles des sujets infectés, le VHE se présente comme une particule sphérique de 27–34 nm de diamètre à symétrie icosaédrique. Le génome viral est un ARN de polarité positive de 7,2 Kb comportant deux régions non codantes en 5’ et en 3’ et 3 cadres ouverts de lecture (ORF) (Fig. 1). L’ORF-1 code une polyprotéine non structurale contenant notamment un domaine ARN polymérase ARN-dépendante. L’ORF-2 code la protéine de capside, cible des anticorps neutralisants ; les propriétés d’auto-assemblage et de conservation antigénique de la protéine tronquée exprimée dans des systèmes recombinants sont la base du développement vaccinal [6]. L’ORF-3, chevauche l’ORF-2 et code une petite phosphoprotéine essentielle à la morphogenèse et la libération des virions. En culture cellulaire et dans le sang circulant, les particules virales sont associées à des lipides dérivés des exosomes et à cette phosphoprotéine, et sont résistantes à la neutralisation par des anticorps [7]. Comme pour le virus de l’hépatite C, la réplication est cytoplasmique et il n’existe pas de forme de latence.
Figure 1. Organisation génomique du VHE
Le VHE a longtemps été le seul membre du genre hepevirus et de la famille des hepeviridae. L’identification de nouveaux virus VHE-like dans un nombre croissant d’espèces animales a conduit le comité international de taxonomie des virus (ICTV) à revoir régulièrement la classification (historique disponible sur le site http://www.ictvonline.org/taxonomyHistory.asp?taxnode_id=20142429&taxa_name=Hepeviridae). Depuis 2015, la famille des Hepeviridae est ainsi divisée en 2 genres : les Orthohepevirus, incluant la plupart des virus infectant les mammifères, les virus infectant les chauves-souris et les virus aviaires, et les Piscihepevirus actuellement retrouvés chez la truite. Les Orthohepevirus sont ensuite divisés en quatre espèces, de A à D. L’espèce A Orthohepevirus regroupe des souches divergentes isolées chez le sanglier et le chameau ainsi que les virus de génotype 1 à 4 retrouvés chez l’homme, classiquement définis par une divergence supérieure à 20 % de la séquence nucléotidique dans la région ORF-2 (Fig. 2) [8].
Figure 2. Analyse phylogénétique des membres de l’espèce des Orthohepevirus A. D’après (Smith et al., 2014)
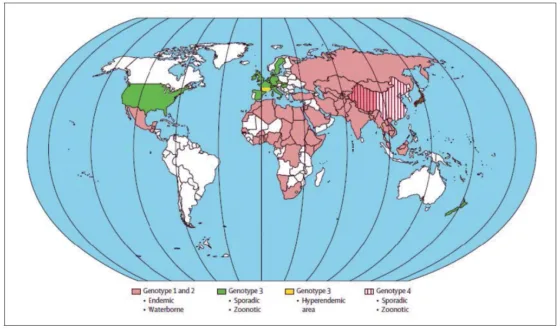
Malgré cette divergence nucléotidique, ces quatre génotypes présentent une communauté antigénique avec un seul sérotype décrit [9]. Les génotypes 1 et 2 sont exclusivement retrouvés chez l’homme, essentiellement dans des zones endémiques telles que le sous-continent indien et l’Afrique. Les génotypes 3 et 4 sont zoonotiques et sont retrouvés à la fois chez l’homme et le porc mais aussi chez le sanglier, les cervidés, le lapin, et probablement d’autres mammifères. Le génotype 3 est responsable de la plupart des infections autochtones en Europe et aux USA alors que le génotype 4 est retrouvé en Asie (Fig. 3) [10].
Figure 3. Distribution géographique génotypes responsable des infections VHE. (en blanc : données insuffisantes, mais plusieurs cas d’infection à génotype 3 ont été rapportés en Amérique du Sud) ; d’après (Kamar et al., 2012a)
La sous-division des quatre génotypes infectant l’homme en de nombreux sous-types [11] a montré le regroupement géographique de ces sous-types. Ainsi les souches les plus fréquemment rencontrées en Europe et en France étaient dans l’ordre décroissant de sous-type 3f, 3c et 3e, avec la même répartition dans les élevages porcins [12].
Transmission et prévention du VHE
Les modes de transmission du VHE varient selon la zone géographique et le génotype viral et conditionnent les actions de prévention (Fig. 4).
Figure 4. Sources et modes de transmission des VHE de génotype 1 à 4 ; d’après (Kamar, 2012 #19)
Dans les pays en développement, les virus de génotype 1 et 2, strictement humains, sont transmis par voie féco-orale via l’ingestion d’eau contaminée. Le réservoir environnemental, alimenté par l’excrétion fécale des sujets infectés, symptomatiques ou non, joue ainsi un rôle majeur dans le maintien de l’endémicité et la survenue d’épidémies [10]. La transmission verticale ou périnatale s’observe exclusivement dans les pays en développement avec des souches de génotype 1. Elle peut s’accompagner d’une mortinatalité importante [10]. La prévention de ces infections passe par l’amélioration des conditions sanitaires et l’accès à l’eau potable et des mesures d’hygiène universelle. Selon un rapport de l’OMS, près de 2 milliards et demi d’individus n’auront toujours pas accès à un réseau d’assainissement en 2015 (Improving nutrition outcomes with better water, sanitation and hygiene ; http://apps.who.int/iris/bitstream/10665/193991/1/9789241565103_eng.pdf).
Dans les pays développés, le réservoir animal, essentiellement porcin, joue un rôle majeur. La transmission alimentaire par ingestion de viande crue ou insuffisamment cuite a été démontrée par homologie des souches entre l’aliment incriminé et le sujet infecté et par des études cas-contrôle. La figatelle corse, saucisse fumée à base de foie de porc, a ainsi été incriminée à plusieurs reprises en France. En Europe, au Japon mais également aux USA, l’ARN viral est détecté dans les foies de porcs entrant dans la chaîne alimentaire à des fréquences atteignant 11 % [13]. Ce mode de transmission peut être évité par la cuisson « cuite à cœur » de la viande porcine et du gibier chez les personnes à risque de développer une forme grave (ces précautions sont résumées dans une affichette destinée aux professionnels de santé sur le site du ministère (http://www.sante.gouv.fr/IMG/pdf/Fiche_Hepatite_E.pdf). En effet, le VHE reste infectieux après 1 h à 56 °C et une cuisson de 20 mn à 71 °C est nécessaire à son inactivation [14].
La contamination de l’environnement par les souches zoonotiques pourrait également jouer un rôle dans la contamination alimentaire des pays développés. En effet, l’ARN viral est détectable dans le lisier et dans les eaux de surface, et dans des aliments tels que les salades ou les framboises de plein champ ou encore les coquillages : au Royaume Uni, du VHE de génotype 3 a été retrouvé dans 39 % des moules recueillies à proximité d’un élevage porcin [13]. Ce mode de transmission est encore mal évalué et rend sa prévention difficile.
Le contact direct avec le réservoir animal est également source d’infection comme le montrent les séroprévalences élevées chez les personnels exposés, vétérinaires, éleveurs, bouchers, chasseurs… Ce mode de transmission peut être prévenu par des mesures d’hygiène simples, telles que le port de gants par les personnels exposés ou le port de bottes en forêt [13].
La transmission transfusionnelle a été démontrée dans de nombreux pays occidentaux, notamment en France, et tous les produits sanguins labiles ont été incriminés. Le risque de don virémique varie selon les pays mais est particulièrement élevé en France, à 1 sur 2218 [15]. Toutefois, l’impact de cette contamination en termes de santé publique reste débattu. Ainsi, au Royaume Uni « seulement » 42 % des receveurs de produits contaminés ont été infectés, sans morbidité significative [16]. En France, depuis novembre 2014 il est possible d’obtenir du plasma VHE négatif, l’ARN viral étant recherché dans une fraction des plasmas sécurisés (quarantaine) ou traités à l’amotosalen.
Diagnostic et données de prévalence
Après une période d’incubation de 3 à 6 semaines, la cytolyse hépatique s’accompagne de l’apparition d’IgM anti-VHE qui peuvent rester détectables plusieurs mois (Fig. 5). Les IgG apparaissent peu de temps après et persistent plusieurs années, mais des réversions sont possibles. L’ARN viral est détectable dans le sang et les selles avant la phase aiguë. En cas d’infection aiguë spontanément résolutive, la virémie se négative en 2 à 3 semaines, parallèlement à la normalisation des transaminases [10]. La virémie peut déjà être négative si l’infection est vue tardivement.
Figure 5. Cinétique des marqueurs virologiques lors de l’infection VHE ; d’après (Kamar et al., 2012a)
Le diagnostic de l’infection aiguë repose sur la détection d’IgM anti-VHE. Les trousses actuellement disponibles en France, tests ELISA ou tests rapides, on une bonne sensibilité et une bonne spécificité chez l’immunocompétent. Comme pour l’hépatite A, des séroconversions retardées peuvent être observées et il ne faut pas hésiter à répéter les tests si une cytolyse reste inexpliquée. Les performances des tests sont moins bonnes chez l’immunodéprimé où la recherche d’IgM doit être associée à la recherche de l’ARN viral, dont la persistance au-delà de 3 à 6 mois permet de définir l’infection chronique. La performance des tests moléculaires s’est améliorée considérablement ces dernières années, notamment grâce au développement de standards internationaux permettant de déterminer leur limite de détection mais également leur sensibilité vis-à-vis des différents génotypes. Plusieurs de ces trousses ont obtenu le marquage CE, dont les trousses Ceeram et Altona.
L’algorithme diagnostique proposé par le Centre National de Référence des Virus des hépatites à transmission entérique est présenté figure 6.
Figure 6. Algorithme diagnostique de l’infection VHE
L’amélioration des performances des tests, puis leur diffusion à un nombre croissant de laboratoires, rend le diagnostic de l’infection accessible à toutes les structures. Cela, ajouté à une meilleure information des praticiens, a conduit à l’identification de plus de 1800 cas d’infection VHE en France en 2014 parmi plus de 40000 patients testés, avec un pourcentage d’infections parmi les patients testés, chiffre stable depuis 2012 autour de 4–5 % ; les infections importées représentent moins de 1 % des cas diagnostiqués (données CNR, disponibles sur http://www.cnrvha-vhe.org/wp-content/uploads/2012/03/2014-Rap-Act-VHE-VHA.pdf).
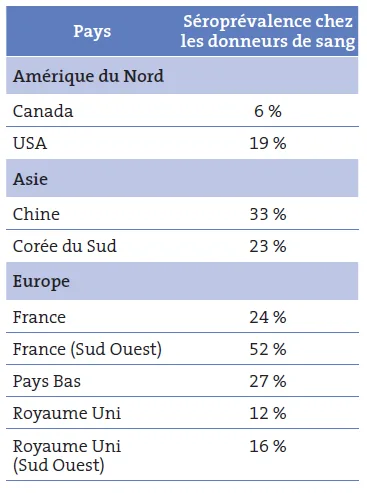
Tableau I. Séroprévalence du VHE chez les donneurs de sang (test IgG Wantai) ; d’après (Petrik, 2015)
La présence d’IgG anti-VHE est le marqueur d’une infection ancienne. La sensibilité des trousses disponibles varie de 0,25 unités OMS/ml pour la trousse Wantaï à 2,5 unités OMS/ml pour d’autres trousses. Les données de séroprévalence ne peuvent donc être comparées que si un même test est utilisé. Il existe des différences considérables de séroprévalence d’un pays à l’autre et d’une région à l’autre au sein d’un même pays comme illustré tableau I et figure 7, chez les donneurs de sang [17]. Ces fortes données de prévalence soulignent le caractère le plus souvent asymptomatique de l’infection.
Figure 7. Séroprévalence du VHE chez les donneurs de sang en France (Gallian, in press)
Présentation clinique
L’hépatite E est actuellement la première cause d’hépatite aiguë virale dans le monde, y compris dans les pays industrialisés.
Dans les pays en développement où les génotypes 1 et 2 sont endémiques, le VHE est responsable d’infections aiguës symptomatiques chez les jeunes adultes (15–30 ans), l’infection étant asymptomatique dans plus de 80 % des cas. Les prodromes et les symptômes ne diffèrent pas des autres formes d’hépatites. Un ictère est présent dans près de la moitié des cas. Des formes sévères sont observées chez les patients ayant une hépatopathie sous-jacente et chez les femmes enceintes où la mortalité peut atteindre 25 % au 3e trimestre de la grossesse [10]. Le nombre annuel d’infections symptomatiques a été estimé en 2005 à près de 3 millions dont 70000 décès et 3000 mortinaissances, correspondant à un taux de mortalité compris entre 0.5 et 4 % [18]. La physiopathologie de l’hépatite fulminante chez la femme enceinte reste mystérieuse et n’est décrite que pour les infections à génotype 1.
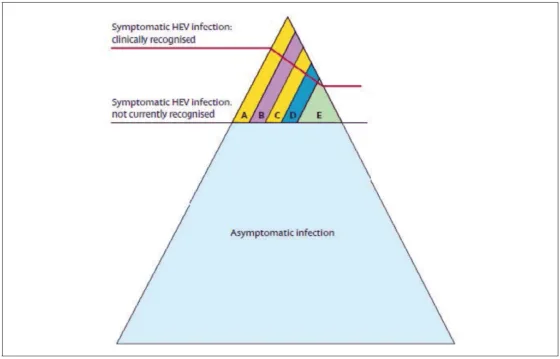
Dans les pays développés, où les génotypes zoonotiques 3 et 4 sont endémiques, l’infection est le plus souvent asymptomatique. L’infection symptomatique est observée chez des patients d’âge moyen (âge médian 50–55 ans), plus fréquemment des hommes, qui présentent des comorbidités ou une consommation excessive d’alcool. La fréquence de l’infection justifie la réalisation d’une sérologie du VHE en première intention devant tout tableau d’hépatite aiguë, a fortiori si celle-ci survient après 40 ans. L’infection VHE peut être source de décompensation de cirrhose ou révéler une hépatopathie sous-jacente. Elle peut également être confondue ou s’associer à une toxicité médicamenteuse [9]. Les infections à génotype 4 ont été décrites comme plus sévères (ALAT plus élevées et ictère plus fréquent) et avec un sex ratio moins marqué que les infections à génotype 3, mais ce génotype reste rarement isolé en France [19]. Les points remarquables des infections à génotype 3 sont la survenue de manifestations extra-hépatiques et le développement d’infections chroniques. L’éventail des présentations cliniques associées aux infections VHE zoonotiques est présenté figure 8.
Figure 8. La plupart des infections VHE de génotype 3 et 4 sont asymptomatiques ou non diagnostiquées. Quand elles sont diagnostiquées (A) Infection aiguë ictérique (B) Infection chronique (C) Contexte de toxicité médicamenteuse (D) Manifestations neurologiques (E) Autres manifestations extra-hépatiques ; d’après (Kamar et al., 2012a)
Les troubles neurologiques sont les manifestations extra-hépatiques les plus fréquemment associées à l’infection VHE de génotype 1 et 3. Il s’agit de syndrome de Guillain Barré, de méningo-encéphalite, de myélite aiguë transverse, ou de syndrome de Parsonage-Turner (amyotrophie névralgique de l’épaule). Ces troubles sont rapportés dans 5 à 10 % des infections VHE de génotype 3, aussi bien aiguës que chroniques. L’ARN viral peut être détectable dans le liquide céphalo-rachidien. Les autres manifestations extra-hépatiques incluent des atteintes rénales (glomérulonéphrites extramembraneuses, membranoprolifératives ou cryoglobulinémiques), des troubles hématologiques ou des atteintes pancréatiques [10].
L’infection chronique est décrite pour les infections de génotype 3 et n’est à ce jour rapportée que chez des sujets immunodéprimés, bien qu’un cas d’infection persistante ait été décrit chez un sujet lupique en rémission [20]. Ces infections persistantes sont décrites essentiellement en Europe chez des transplantés d’organe solide (rein, foie, cœur) où le risque de passage à la chronicité atteint 66 %. Chez ces patients, la stabilité de la charge virale et l’absence de clearance virale à 3 mois de l’infection permet d’établir son caractère chronique [21]. Cette infection est asymptomatique dans près de 70 % des cas et s’accompagne d’une élévation modérée des transaminases, mais la progression de la fibrose peut être particulièrement rapide. Des études transversales ont montré que l’ARN viral était détectable chez 0.9–3.5 % des transplantés et que cette fréquence passait à 4.3–6.5 % quand des transplantés à transaminases élevées étaient ciblés [10]. L’ensemble de ces éléments, notamment le caractère asymptomatique de l’infection, fait recommander une recherche systématique de l’ARN viral chez tout patient transplanté ; en effet le diagnostic sérologique peut être mis en défaut dans ce contexte avec une séroconversion anti-VHE retardée ou absente. D’autres immunodéprimés sont concernés par l’infection VHE chronique, tels les patients vivant avec le VIH et ayant un taux de CD4 < 250/mm3, les greffés de moelle et les patients sous chimiothérapie ou immunothérapie. Les modes de contamination retrouvés chez les immunodéprimés sont similaires à ceux de la population générale, essentiellement consommation de porc, de gibier ou de coquillages, mais des cas de transmission transfusionnelle ou par le greffon ont été décrits [10].
Traitement de l’infection
L’infection VHE chronique est le plus souvent associée à un traitement immunosuppresseur qui inhibe l’activation des cellules T et compromet la réponse anti-VHE. Cette réponse spécifique est altérée lors de l’infection chronique et est restaurée après traitement [22]. Quand elle est possible, la réduction de l’immunosuppression permet l’élimination du VHE chez près de 30 % des sujets transplantés [23]. En effet, outre la suppression de la réponse immunitaire nécessaire au contrôle de l’infection virale, certains immunosuppresseurs ciblant les cellules T, tels les anti-calcineurines ou les inhibiteurs de mTOR, stimulent la réplication du VHE in vitro [22]. Cette approche ne peut être entreprise que chez les greffés relevant d’une immunosuppression faible (greffés hépatiques par exemple).
Si cette stratégie est impossible ou inefficace, l’interféron alpha pégylé ou la ribavirine peuvent être utilisés. Toutefois, l’utilisation de l’interféron est limitée dans le contexte de greffe d’organe en raison du risque de rejet. La ribavirine est alors le traitement de choix. Prescrite à la dose de 10–12 mg/kg par jour pendant 3 mois, elle permet l’élimination virale dans près de 80 % des cas chez les transplantés d’organe [24]. Elle agit de manière indirecte sur la réplication virale par déplétion du pool cellulaire de guanosine tri-phosphate [25]. Les échecs de ce traitement sont souvent associés à une réduction de dose opérée devant la survenue d’anémie sévère. Il n’existe pas de résistance génotypique à la ribavirine, mais une mutation augmentant la capacité réplicative du virus a récemment été identifiée dans la région codant l’ARN polymérase virale (G1634R). Cette mutation préexiste au traitement et s’accompagne de charges virales plus élevées, mais n’impacte pas la réponse virologique (Lhomme, in press).
Les sujets immunocompétents éliminent spontanément le virus sans nécessité de traitement antiviral. Quelques auteurs ont néanmoins rapporté l’utilisation de la ribavirine dans des formes aiguës fulminantes ou chez des sujets risquant de décompenser une hépatopathie sous-jacente [10]. L’absence de bras contrôle, difficile a obtenir dans cette population, ne permet pas d’établir le réel bénéfice de cette molécule. De plus, la ribavirine étant tératogène, elle n’est pas utilisable chez la femme enceinte. Des nouvelles stratégies antivirales sont donc nécessaires. Ainsi, le sofosbuvir, analogue nucléotidique actif sur l’ARN polymérase du virus de l’hépatite C, a montré une activité anti-VHE in vitro et pourrait donc présenter une alternative à la ribavirine [26].
Prévention de l’infection par le VHE
Outre les mesures rappelées au chapitre « modes de transmission et prévention », beaucoup d’espoirs résident dans la vaccination. En l’absence de système de culture efficace, des vaccins anti-VHE ont pu être développés grâce aux propriétés d’auto-assemblage de peptides issus de la région ORF-2 codant la capside virale. Ces pseudo-particules ont des caractéristiques antigéniques similaires au virion natif. Un seul vaccin, basé sur un peptide recombinant de 239 acides aminés (Hecolin), est actuellement homologué et commercialisé en Chine depuis 2011 pour les sujets de plus de 16 ans. L’injection de 3 doses sur 6 mois induit une séroconversion chez pratiquement tous les sujets. Les données des essais cliniques réalisés en Chine font état d’une efficacité vaccinale de 95 % à 12 mois [27] et de 87 % jusqu’à 4,5 ans après vaccination et une protection essentiellement contre les infections de génotype 4, endémique en Chine. Il n’y a pas de données sur son efficacité dans les contextes épidémiques, ni chez la femme enceinte. L’OMS recommande des études de phase 4 pour mieux établir son profil de tolérance et son efficacité dans d’autres contextes (note de synthèse disponible sur le site de l’OMS : http://www.who.int/wer/2015/wer9018.pdf).
Références
Khuroo MS. Study of an epidemic of non-A, non-B hepatitis. Possibility of another human hepatitis virus distinct from post-transfusion non-A, non-B type. Am J Med 1980;68: 818-24.
Balayan MS, Andjaparidze AG, Savinskaya SS, et al. Evidence for a virus in non-A, non-B hepatitis transmitted via the fecal-oral route. Intervirology 1983;20: 23-31.
Reyes GR, Purdy MA, Kim JP, et al. Isolation of a cDNA from the virus responsible for enterically transmitted non-A, non-B hepatitis. Science 1990;247:1335-9.
Meng XJ, Purcell RH, Halbur PG, et al. A novel virus in swine is closely related to the human hepatitis E virus. Proc Natl Acad Sci U S A 1997;94: 9860-5.
Kwo PY, Schlauder GG, Carpenter HA, et al. Acute hepatitis E by a new isolate acquired in the United States. Mayo Clin Proc 1997;72: 1133-6.
Ahmad I, Holla RP, Jameel S. Molecular virology of hepatitis E virus. Virus Res 2011;161: 47-58.
Feng Z, and Lemon SM. Peek-a-boo: membrane hijacking and the pathogenesis of viral hepatitis. Trends Microbiol 2014;22:59-64.
Smith DB, Simmonds P, Jameel S, et al. Consensus proposals for classification of the family Hepeviridae. J Gen Virol 2014; 95(Pt 10):2223-32.
Kamar N, Bendall R, Legrand-Abravanel F, et al. Hepatitis E. Lancet 2012;379:2477-88.
Kamar N, Dalton HR, Abravanel F, et al. Hepatitis E virus infection. Clin Microbiol Rev 2014;27:116-38.
Lu L, Li C, Hagedorn CH. Phylogenetic analysis of global hepatitis E virus sequences: genetic diversity, subtypes and zoonosis. Rev Med Virol 2006;16:5-36.
Bouquet J, Tesse S, Lunazzi A, et al. Close similarity between sequences of hepatitis E virus recovered from humans and swine, France, 2008-2009. Emerg Infect Dis 2011;17: 2018-25.
Pavio N, Meng XJ, Doceul V. Zoonotic origin of hepatitis E. Curr Opin Virol 2015;10:34-41.
Barnaud E, Rogee S, Garry P, et al. Thermal inactivation of infectious hepatitis E virus in experimentally contaminated food. Appl Environ Microbiol 2012;78:5153-9.
Gallian P, Lhomme S, Piquet Y, et al. Hepatitis E virus infections in blood donors, France. Emerg Infect Dis 2014;20:1914-7.
Hewitt PE, Ijaz S, Brailsford SR, et al. Hepatitis E virus in blood components: a prevalence and transmission study in southeast England. Lancet 2014; 384:1766-73.
Petrik J, Lozano M, Seed CR, et al. Hepatitis E. Vox Sanguinis 2015
Rein DB, Stevens GA, Theaker J, et al. The global burden of hepatitis E virus genotypes 1 and 2 in 2005. Hepatology 2012;55: 988-97.
Jeblaoui A, Haim-Boukobza S, Marchadier E, et al. Genotype 4 hepatitis e virus in France: an autochthonous infection with a more severe presentation. Clin Infect Dis 2013;57: e122-6.
Grewal P, Kamili S, Motamed D. Chronic hepatitis E in an immunocompetent patient: a case report. Hepatology 2014;59:347-8.
Kamar N, Garrouste C, Haagsma EB, et al. Factors associated with chronic hepatitis in patients with hepatitis E virus infection who have received solid organ transplants. Gastroenterology 2012;140:1481-9.
Wang Y, Metselaar H J, Peppelenbosch MP, et al. Chronic hepatitis E in solid-organ transplantation: the key implications of immunosuppressants. Curr Opin Infect Dis 2014;27: 303-8.
Kamar N, Garrouste C, Haagsma EB, et al. Factors associated with chronic hepatitis in patients with hepatitis E virus infection who have received solid organ transplants. Gastroenterology 2011;140:1481-9.
Kamar N, Izopet J, Tripon S, et al. Ribavirin for chronic hepatitis E virus infection in transplant recipients. N Engl J Med 2014;370: 1111-20.
Sayed IM, Vercouter AS, Abdelwahab SF, et al. Is hepatitis E virus an emerging problem in industrialized countries? Hepatology 2015;62: 1883-92.
Dao Thi VL, Debing Y, Wu X, et al. Sofosbuvir Inhibits Hepatitis E Virus Replication In Vitro and Results in an Additive Effect When Combined With Ribavirin. Gastroenterology 2016;150:82-85 e4.
Zhu FC, Zhang J, Zhang XF, et al. Efficacy and safety of a recombinant hepatitis E vaccine in healthy adults: a large-scale, randomised, double-blind placebo-controlled, phase 3 trial. Lancet 2010;376:895-902.
Les Cinq points forts
L’infection par le virus E se caractérise par deux profils épidémiologiques distincts correspondant d’une part à des infections par des virus strictement humains relevant du péril fécal dans les pays pauvres et à des infections par des virus zoonotiques dont le réservoir majeur est l’élevage porcin, dans les pays riches.
L’hépatite E est la première cause d’hépatite virale aiguë ; Elle doit être recherchée de façon systématique devant toute hépatite aiguë , y compris après transfusion. Les IgM anti-VHE sont la prescription de première intention chez l’immunocompétent et doivent être systématiquement associées à la recherche de l’ARN viral chez le sujet immunodéprimé.
Le passage à la chronicité ne survient que chez les sujets immuno-déprimés, elle atteint 66 % chez les transplantés d’organe. Une recherche systématique du VHE doit être envisagée, dans cette population, en cas de perturbation des tests hépatiques, pour dépister et traiter une infection qui peut être rapidement fibrosante.
Le traitement de l’infection chronique repose d’abord sur la réduction de l’immunosuppression ; puis en cas d’échec ou d’impossibilité, sur la ribavirine.
La prévention de l’infection VHE dans les pays riches repose sur des mesures d’hygiène de base : cuisson à cœur de la viande de porc et de gibier pour limiter la transmission alimentaire et port de gants et de bottes pour les professionnels à risque pour limiter la transmission par contact.
Toute reproduction ou réécriture, totale ou partielle, sans l’accord préalable écrit de la FMC HGE est interdite.
FMC HGE : Organisme certifié Qualiopi pour la catégorie ACTIONS DE FORMATION.